

Jonges M, Meijer A, Fouchier RA, Koch G, Li J, Pan. Guiding outbreak management by the use of influenza A(H7Nx) virus sequence analysis. Euro Surveill. 2013;18(16):pii=20460.
The recently identified human infections with avian influenza A(H7N9) viruses in China raise important questions regarding possible source and risk to humans. Sequence comparison with an influenza A(H7N7) outbreak in the Netherlands in 2003 and an A(H7N1) epidemic in Italy in 1999–2000 suggests that widespread circulation of A(H7N9) viruses must have occurred in China. The emergence of human adaptation marker PB2 E627K in human A(H7N9) cases parallels that of the fatal A(H7N7) human case in the Netherlands.
Background
Since 31 March 2013, Chinese health authorities have been reporting human cases of avian influenza A(H7N9) virus infections. This novel reassortant influenza virus, carrying six internal gene segments of poultry A(H9N2) viruses, supplemented with a haemagglutinin (HA) subtype 7 and a neuraminidase (NA) subtype 9 originating from wild birds [1,2], has caused infections in at least 82 persons, of whom 17 have died, as of 17 April 2013. The human infections occurred in eastern China in four provinces (Henan, Anhui, Jiangsu, and Zhejiang) and two municipalities (Shanghai and Beijing). Currently, the source of the human infections is unclear. However, in response to the detection of the influenza A(H7N9) virus among chickens, pigeons, ducks and environmental samples from some animal markets, as reported to the World Organisation for Animal Health (OIE), Chinese authorities have suspended live poultry trade and implemented the immediate closure of poultry markets, launched road inspections for transport of poultry, and have culled birds in an effort to deal with the issue. The outbreak raises important questions regarding possible source and risk to humans, and these will be addressed through case investigations. Here, we compare some findings from the first two weeks of the outbreak with those from a large highly pathogenic avian influenza (HPAI) A(H7N7) virus outbreak in the Netherlands in 2003 and from a low pathogenic avian influenza (LPAI) A(H7N1) epidemic in Italy in 1999–2000 [3-5] and discuss issues related to diagnosis and the use of molecular surveillance to monitor the outbreak.
Influenza A(H7N7) outbreak in the Netherlands in 2003
Exactly 10 years ago, the Netherlands was struck by an HPAI A(H7N7) virus outbreak that resulted in the infection of poultry on 255 farms and the subsequent culling of about 30 million chickens. A total of 453 exposed persons had mild symptoms and were investigated, of whom 89 were laboratory-confirmed as having an A(H7N7) virus infection [6,7].
Diagnosis of influenza A(H7Nx) virus infection
During the HPAI A(H7N7) virus outbreak in the Netherlands, almost all human cases had mild symptoms, particularly conjunctivitis, but one veterinarian died after an episode of severe influenza-like illness complicated by acute respiratory distress syndrome (ARDS) [7] . Diagnosis was based on virus detection by reverse transcription polymerase chain reaction (RT-PCR) from eye swabs, or combined nose and throat swabs. An important observation was that the sensitivity of eye swab-based diagnostics was much higher than that of diagnostics based on combined nose and throat swabs [6,7]. Similarly, in later sporadic infections of humans with H7 influenza A viruses, ocular symptoms were observed, probably caused by a preference of H7 influenza viruses for receptors in the eye [8]. Studies have shown that H7 influenza viruses may use the ocular mucosa as portal of entry for systemic infection and that this is strain dependent [9,10]. Such symptoms have not been described for the cases of A(H7N9) virus infection in China in 2013, but it may be important to actively monitor for conjunctivitis in the outbreak investigation, as it may increase the success of case finding, particularly for mild cases.
Serological surveillance is important to rule out infection in patients sampled too late for direct virus detection and to assess the extent of transmission. This may be a problem since serological responses in persons with confirmed influenza A(H7Nx) virus infection have been difficult to detect, making assessment of A(H7N9) virus exposure using serosurveys challenging [11,12]. However, determining the kinetics of the antibody response in confirmed cases of influenza A(H7N9) virus infection will provide important information that can inform public health action.
Comparative analysis based on virus sequencing
Detecting the novel virus in animals is challenging as the A(H7N9) virus is a LPAI virus that is expected to cause few or no signs of disease in poultry, allowing silent spread among poultry flocks. The sharing of influenza A(H7N9) virus sequence data by both Chinese veterinary and public health institutes through the Global Initiative on Sharing All Influenza Data (GISAID) allows comparison with the sequences obtained during the Dutch outbreak. We therefore performed a comparative analysis using HA, NA and PB2 (subunit of the influenza virus RNA polymerase complex) fragment sequences from Chinese A(H7N9) viruses in 2013, Dutch A(H7N7) viruses in 2003 and sequences from a well-described LPAI A(H7N1) epidemic in Italy in 1999–2000 [5]. Providers of sequences downloaded from GISAID, listed with accession numbers, are acknowledged in the Table.
?
Table. Origin of the sequences of influenza A(H7Nx) viruses used for the comparative analysis
Sequence analysis of the Dutch viruses detected in poultry and in humans showed rapid diversification of the outbreak strain into multiple lineages (Figure). On the basis of the combined epidemiological and laboratory analyses, we demonstrated that sequences from humans were positioned mostly at ends of the branches of minimal spanning trees, confirming that humans were probably not involved in onward transmission [3].
Figure. Genetic diversity of three influenza A(H7Nx) virus outbreaks expressed by minimum spanning trees 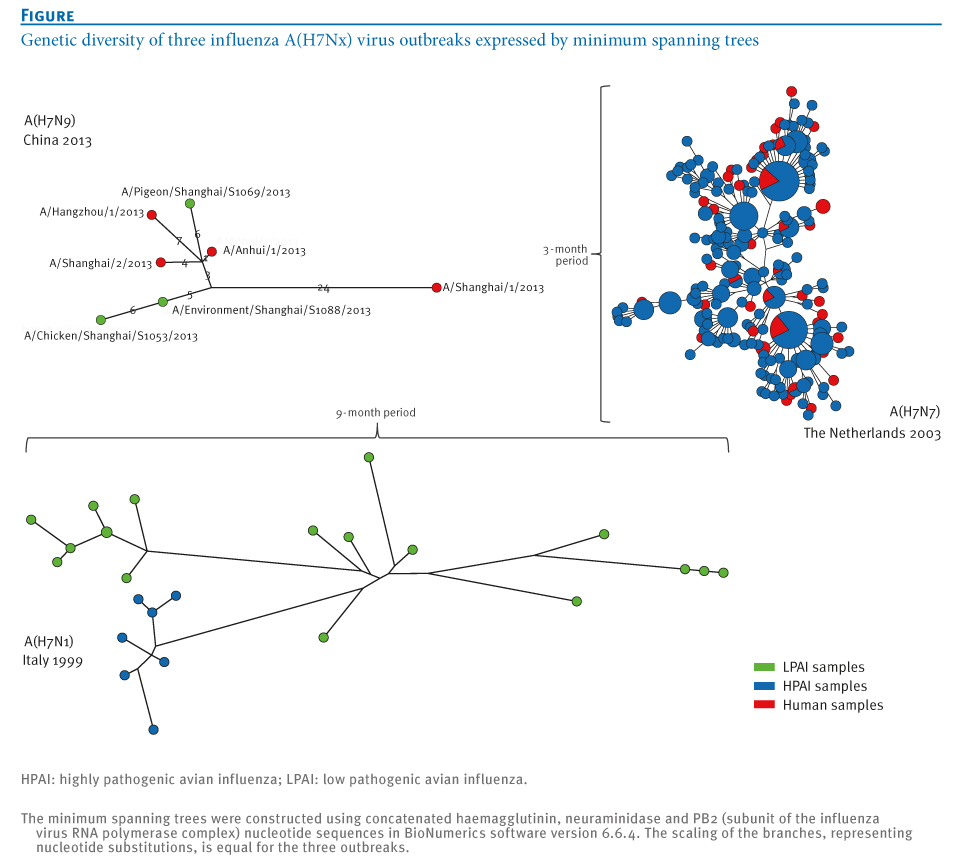
In the current study, we compared the sequence diversity observed during the Dutch A(H7N7) outbreak and Italian A(H7N1) epidemic with the initial A(H7N9) virus sequences from the current outbreak in China. The maximum genetic distance generated during the three months of the Dutch HPAI A(H7N7) outbreak in concatenated HA, NA and PB2 segments of A(H7N7) viruses was 25 nucleotide substitutions. For the Italian LPAI A(H7N1) epidemic, the distance generated during a nine-month period was 66 nucleotide substitutions. For the A(H7N9) outbreak strains, this genetic distance is 35 substitutions, or 21 substitutions when the outlier strain A/Shanghai/1/2013 is ignored (Figure).
All (n=7) NA sequences of the A(H7N9) viruses are characterised by a deletion in the stalk region, associated with adaptation to gallinaceous hosts [1,2,13]. Similar deletions in the NA stalk were also observed during the A(H7N7) outbreak in the Netherlands and the A(H7N1) epidemic in Italy [5]. Given the degree of sequence diversity present in initial A(H7N9) virus sequences, compared with that of the Dutch HPAI A(H7N7) and Italian LPAI A(H7N1) outbreak strains, and the large geographical area affected, the data are suggestive of (silent) spread and adaptation in domestic animals before the novel A(H7N9) virus was identified in humans.
Human adaptation markers
The majority of the Dutch human cases of A(H7N7) virus infection had mild symptoms, with the exception of one fatal case who was diagnosed with an A(H7N7) virus with the mammalian adaptation marker PB2 E627K. This mutation most probably occurred during infection of this case and was associated with high virulence [14]. Remarkably, the PB2 segments of the four available human virus genome sequences from China all carry this E627K substitution, which is absent in the virus isolates obtained from birds and the environment [2]. In addition, three of the four infections with the virus with PB2 E627K were fatal. There are two plausible explanations for this observation:
- the mammalian adaptation markers are selected during replication in humans following exposure to viruses that do not have this mutation, which are circulating in animals;
- the mammalian adaptation markers result from virus replication in animals from which humans become infected.
The relatively protracted disease course in the current outbreak of A(H7N9) virus infection, with relatively mild symptoms at first, followed by exacerbation in the course of a week or longer, is suggestive of the first hypothesis, similar to the outbreak in the Netherlands. In this scenario, an important difference in the A(H7N7) observations from the Netherlands is the frequency of finding the PB2 E627K mutation in humans (4/4 A(H7N9) sequenced patient strains compared with 1/61 sequenced A(H7N7) patient strains). Therefore, an outstanding question is whether the A(H7N9) viruses are more readily mutating in humans or milder cases are being missed. Contact investigations have found no mild cases and only one asymptomatic case), but in order to address this issue, more enhanced testing of persons exposed to a similar source is needed, using the most sensitive tests available on the optimal clinical specimen type obtained at the right time.
Although human infections with H7 influenza viruses have occurred repeatedly over the last decades without evidence of sustained human-to-human transmission, the absence of sustained human-to-human transmission of A(H7N9) viruses does not come with any guarantee. Five of seven A(H7N9) virus strains obtained from humans (n=2), birds (n=2) and the environment (n=1) have a mutation in HA, Q226L, that is associated with binding to alpha(2,6)-linked sialic acids, the virus receptors in the human upper respiratory tract [2]. This Q226L substitution in combination with G228S has been associated with human receptor preference for influenza viruses that caused the pandemics of 1957 and 1968 and with airborne transmission of A(H5N1) virus [15,16]. For H7 viruses, it has recently been demonstrated that these mutations also increased human receptor-binding affinity [17]. In combination with the PB2 E627K mutation, the A(H7N9) virus thus contains two well-known mammalian adaptation markers.
Conclusion
Comparative analysis of the first virological findings from the current outbreak of influenza A(H7N9) virus infection in China with those from other influenza A(H7Nx) outbreaks suggests that widespread circulation must have occurred, resulting in major genetic diversification. Such diversification is of concern, given that several markers associated with increased risk for public health are already present. Enhanced monitoring of avian and mammalian animal reservoirs is of utmost importance as the public health risk of these A(H7N9) viruses may change following limited additional modification.
Acknowledgements
This work was supported by the Dutch Ministry of Economic Affairs, Agriculture, and Innovation, Castellum Project.
We acknowledge the authors, originating and submitting laboratories of the sequences from the Global Initiative on Sharing All Influenza Data (GISAID)s EpiFlu Database (www.gisaid.org), on which this research is based.
Authors’ contributions
Yue-Long Shu, Hualan Chen, Jun Li, Jing-Cao Pan, Ron A.M. Fouchier and Guus Koch improved the manuscript following writing by Marcel Jonges, Adam Meijer and Marion Koopmans. All authors were directly involved in the generation, sharing and analysis of influenza sequence data.
Conflict of interest
None declared.
References
- Gao R, Cao B, Hu Y, Feng Z, Wang D, Hu W, et al. Human infection with a novel avian-origin influenza A (H7N9) virus. N Engl J Med. 2013 Apr 11. [Epub ahead of print].
- Kageyama T, Fujisaki S, Takashita E, Xu H, Yamada S, Uchida Y, Neumann G, et al. Genetic analysis of novel avian A(H7N9) influenza viruses isolated from patients in China, February to April 2013. . Euro Surveill. 2013;18(15):pii=20453. Available from: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=20453
- Jonges M, Bataille A, Enserink R, Meijer A, Fouchier RA, Stegeman A, et al. Comparative analysis of avian influenza virus diversity in poultry and humans during a highly pathogenic avian influenza A (H7N7) virus outbreak. J Virol. 2011;85(20):10598-604.
http://dx.doi.org/10.1128/JVI.05369-11
PMid:21849451 PMCid:3187520 - Bataille A, van der Meer F, Stegeman A, Koch G. Evolutionary analysis of inter-farm transmission dynamics in a highly pathogenic avian influenza epidemic. PLoS Pathog. 2011;7(6):e1002094.
- Capua I, Mutinelli F, Pozza MD, Donatelli I, Puzelli S, Cancellotti FM. The 1999-2000 avian influenza (H7N1) epidemic in Italy: veterinary and human health implications. Acta Trop. 2002;83(1):7-11.
http://dx.doi.org/10.1016/S0001-706X(02)00057-8 - Koopmans M, Wilbrink B, Conyn M, Natrop G, van der Nat H, Vennema H, et al. Transmission of H7N7 avian influenza A virus to human beings during a large outbreak in commercial poultry farms in the Netherlands. Lancet. 2004;363(9409):587-93.
http://dx.doi.org/10.1016/S0140-6736(04)15589-X - Fouchier RA, Schneeberger PM, Rozendaal FW, Broekman JM, Kemink SA, Munster V, et al. Avian influenza A virus (H7N7) associated with human conjunctivitis and a fatal case of acute respiratory distress syndrome. Proc Natl Acad Sci U S A. 2004;101(5):1356-61.
http://dx.doi.org/10.1073/pnas.0308352100
PMid:14745020 PMCid:337057 - Olofsson S, Kumlin U, Dimock K, Arnberg N. Avian influenza and sialic acid receptors: more than meets the eye? Lancet Infect Dis. 2005;5(3):184-8.
- Belser JA, Wadford DA, Xu J, Katz JM, Tumpey TM. Ocular infection of mice with influenza A (H7) viruses: a site of primary replication and spread to the respiratory tract. J Virol. 2009;83(14):7075-84.
http://dx.doi.org/10.1128/JVI.00535-09
PMid:19458003 PMCid:2704783 - Belser JA, Davis CT, Balish A, Edwards LE, Zeng H, Maines TR, et al. Pathogenesis, transmissibility, and ocular tropism of a highly pathogenic avian influenza A (H7N3) virus associated with human conjunctivitis. J Virol. 2013 Mar 13. [Epub ahead of print].
- Meijer A, Bosman A, van de Kamp EE, Wilbrink B, Du Ry van Beest Holle M, Koopmans M. Measurement of antibodies to avian influenza virus A(H7N7) in humans by hemagglutination inhibition test. J Virol Methods. 2006;132(1-2):113-20.
- Min JY, Vogel L, Matsuoka Y, Lu B, Swayne D, Jin H, et al. A live attenuated H7N7 candidate vaccine virus induces neutralizing antibody that confers protection from challenge in mice, ferrets, and monkeys. J Virol. 2010;84(22):11950-60.
http://dx.doi.org/10.1128/JVI.01305-10
PMid:20810733 PMCid:2977864 - Li J, Zu Dohna H, Cardona CJ, Miller J, Carpenter TE. Emergence and genetic variation of neuraminidase stalk deletions in avian influenza viruses. PLoS One. 2011;6(2):e14722.
- de Wit E, Munster VJ, van Riel D, Beyer WE, Rimmelzwaan GF, Kuiken T, et al. Molecular determinants of adaptation of highly pathogenic avian influenza H7N7 viruses to efficient replication in the human host. J Virol. 2010;84(3):1597-606.
http://dx.doi.org/10.1128/JVI.01783-09
PMid:19939933 PMCid:2812334 - Pappas C, Viswanathan K, Chandrasekaran A, Raman R, Katz JM, Sasisekharan R, et al. Receptor specificity and transmission of H2N2 subtype viruses isolated from the pandemic of 1957. PLoS One. 2010;5(6):e11158.
- Herfst S, Schrauwen EJ, Linster M, Chutinimitkul S, de Wit E, Munster VJ, et al. Airborne transmission of influenza A/H5N1 virus between ferrets. Science. 2012;336(6088):1534-41.
http://dx.doi.org/10.1126/science.1213362
PMid:22723413 - Srinivasan K, Raman R, Jayaraman A, Viswanathan K, Sasisekharan R. Quantitative description of glycan-receptor binding of influenza A virus H7 hemagglutinin. PLoS One. 2013;8(2):e49597.
See Also:
Latest articles in those days:
- The surveillance programme for avian influenza (AI) in Norwegian wildlife 2025 6 hours ago
- The surveillance programme for avian influenza (AI) in poultry in Norway 2025 6 hours ago
- Emergence of Novel Reassortant H3N2 Avian Influenza Viruses in Southern China: Genetic Complexity and Pathogenicity in Chickens and Mice 6 hours ago
- Pathological evidence of neurotropism and oculotropism in wild black-headed gulls naturally infected with H5N1 high pathogenicity avian influenza 6 hours ago
- Birth cohort effects in adults associated with influenza A(H1N1)pdm09 vaccine effectiveness 18 hours ago
[Go Top] [Close Window]