

Skowronski DM, De Serres G, Janjua NZ, Gardy JL, G. Cross-reactive antibody to swine influenza A(H3N2) subtype virus in children and adults before and after immunisation with 2010/11 trivalent inactivated influenza vaccine in Canada, August to November. Euro Surveill. 2012;17(4):pii=20066
In pre- and post-immunisation sera from children (17–120 months-old) and adults (20–59 years-old) immunised with 2010/11 trivalent inactivated influenza vaccine, we assessed age-related patterns of sero-susceptibility and vaccine-induced cross-reactive antibodies to a representative swine H3N2 (swH3N2) and a related ancestral human H3N2 (A/Sydney/5/1997) influenza virus. Few children but a greater proportion of adults showed pre-immunisation haemagglutination inhibition titres ≥40 to either virus. Titres increased with age among children but decreased in adults. Fewer than 20% showed a four-fold rise in antibody titres to either virus following immunisation. Further investigation is warranted to guide ongoing risk assessment and response to emerging swine H3N2 viruses.
Introduction
The first documented human infection in North America with an influenza A(H3N2) virus of swine origin (swH3N2) occurred in an Ontario farm worker in 2005 (A/Ontario/RV1273/2005). It involved a swH3N2 lineage that had entered swine from humans in the mid-1990s [1,2]. Sporadic human cases occurred thereafter in the United States (US) and Canada [3,4]. During the latter half of 2011, 12 cases of human infection with a variant of swH3N2 (designated A(H3N2)v by the World Health Organization) [5] were identified in the US, primarily among children including some without recognised swine exposure [6]. Recent analysis has shown that swH3N2 viruses and zoonotic transmissions to humans, including A(H3N2)v, are descendants from a common human influenza virus ancestor, the A/Wuhan/359/1995(H3N2)-like virus [7]. Influenza A/Wuhan/359/1995 has not circulated in humans nor been a component of the trivalent inactivated influenza vaccine (TIV) since 1998, when it was replaced by influenza A/Sydney/5/1997(H3N2) [8-10].
Herein we assess cross-reactive antibody titres to a representative swH3N2 virus in sera collected from Canadian children and adults before and after immunisation with the 2010/11 TIV, containing the same vaccine components as the 2011/12 formulation [10].
Methods
We used a convenience sample of previously collected pre- and post-immunisation sera from children and adults enrolled in 2010/11 TIV immunogenicity trials. Sera had been collected at baseline and 21 to 28 days after the last age-appropriate dose of TIV (Fluviral, GSK, Laval, Quebec, Canada) from Quebec adults in August and September 2010 and from Quebec children in October and November 2010. All had been vaccinated in 2009 with the monovalent AS03-adjuvanted influenza A(H1N1)pdm09 vaccine (Arepanrix; GSK, Laval, Quebec). The study protocols have been described previously [11,12] and had been approved by the ethics board of the Centre Hospitalier Universitaire de Québec.
Sera were tested for antibodies (i) to influenza A/Wisconsin/15/2009(H3N2), considered antigenically equivalent to the influenza A/Perth/16/2009(H3N2)-like component of the 2010/11 (and also 2011/12) northern hemisphere TIV (referred to in this paper as A/Wisconsin), (ii) to a swH3N2 virus (A/ferret/QC/844/2011; F844) isolated from a ferret infected in February 2011 while temporarily housed with swine at the same Quebec animal research facility, and (iii) to influenza A/Sydney/5/97(H3N2) as a human influenza ancestor of swH3N2 (A/Sydney). Because influenza A/Wuhan/359/1995 virus was not readily accessible, A/Sydney was chosen as the most closely related, available alternative against which to compare age-related trends in cross-reactive antibody levels.
Multiple sequence alignments [13] and BLAST searches [14] generated pairwise identities between F844 gene segments (GenBank accession numbers JQ409334 to JQ409341) and available segments from the following viruses available in GISAID: A(H3N2)v (A/Indiana/10/2011(passage X-1) (Indiana State Department of Health Laboratories, Centers for Disease Control and Prevention, sequence authors B Shu, R Garten, S Emery, A Balish, C Smith, J Barnes, S Lindstrom, A Klimov, N Cox) , A/Wisconsin (passage X-183) (Wisconsin State Laboratory of Hygiene, Centers for Disease Control and Prevention, sequence authors not specified), A/Wuhan/359/1995 (imported from NCBI) and A/Sydney (imported from NCBI).
Haemagglutinin (HA) identity was assessed across the HA1 peptide and antigenic regions defined by one scheme of 59 amino acids [7] and an expanded scheme comprising 130 amino acids [15]. Relatedness was further assessed through phylogenies of HA and neuraminidase (NA) surface proteins of F844, A(H3N2)v, other swine and human influenza A(H3N2) isolates, and TIV components [16]. The authors gratefully acknowledge the 268 originating and submitting laboratories who contributed sequences used in the phylogenetic analysis to GISAID, and recognise in particular the labs who contributed swine and A(H3N2)v sequences highlighted in our phylogenetic analysis: Canadian Food Inspection Agency, Centers for Disease Control and Prevention, Indiana State Department of Health Laboratories, Iowa State Hygienic Laboratory, Kansas Department of Health and Environment, Maine Health and Environmental Testing Laboratory, Minnesota Department of Health, Pennsylvania Department of Health, Public Health Agency of Canada, and University of Pittsburgh Medical Center Microbiology Lab.
Antibody titres were measured in duplicate by haemagglutination inhibition (HI) as previously described [11]. Turkey red blood cells (RBCs) were used for F844 and A/Wisconsin; guinea pig RBCs were used for A/Sydney. Titres <10 were assigned a value of 5. Scatter plots and Pearson correlation coefficients of natural logarithm-transformed titres were explored. Immunogenicity end points included group GMTs, the ratio of post- versus pre-immunisation GMTs (GMTR), the proportion of participants with HI titre ≥40 (by convention considered the sero-protective threshold for evaluating vaccine antigens), and the proportion of sero-converting individuals (those showing four-fold increase in post- compared with pre-immunisation titres or from HI titre <10 pre-immunisation to at least 40 post-immunisation) [17,18]. Linear regression models assessed trends in GMTs by one-year age interval and the chi-square test was used to compare differences in the proportion of participants with HI titre ≥40 by age category.
Results
Participants
Sera from 138 children were included. The mean/median age of paediatric participants was 63/63 months, with a range of 17 to 120 months. Forty-six children never before immunised against seasonal influenza received two doses of 2010/11 TIV, whereas 91 received a single dose. For one child this information was not available. Eighty children (58%) had received at least one prior TIV dose, 24 (17%) had received at least three doses. Sixty-five adults were included, among whom the mean/median age was 40/39 years, with a range of 20 to 59 years. Of these, 58 (89%) had received TIV previously: 41 (71%) a single dose, 16 (28%) twice and one (1.7%) three times previously.
Phylogenetic relatedness
Phylogenetic analysis established F844 to be representative of circulating swH3N2 viruses with zoonotic potential, including A(H3N2)v (Figure 1). BLAST indicated the closest match to each F844 segment originated from swH3N2 viruses isolated in North America between 2005 and 2010 (pairwise identities 97.9–98.9%), with the F844 HA and NA most similar to that of the A/swine/QC/382/2009 virus (98.3% and 99.1% pairwise identity, respectively). The closest human swH3N2 HA was found in A/Iowa/16/2009 (97.4% pairwise identity), and the closest human swH3N2 NA in A/Ontario/1252/2007 (98.9% pairwise identity) (Figure 1).
Figure 1. Maximum likelihood phylogeny showing location of F844 relative to swine and human swH3N2 viruses and human vaccine strains 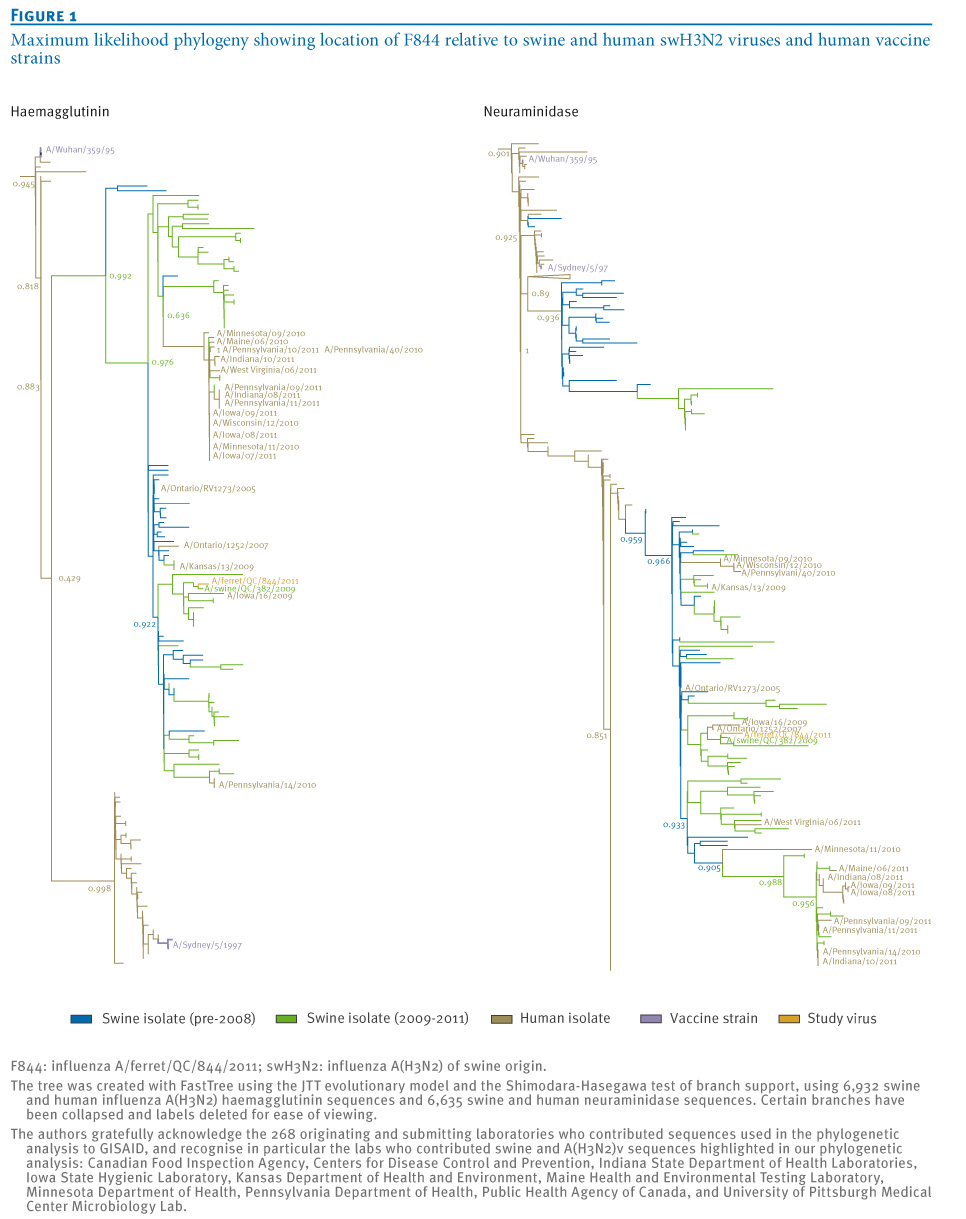
The human-origin isolate influenza A/Ontario/RV1273/2005 was among the ten isolates most closely related to F844 for all but the NS segment, further reinforcing zoonotic potential of the F844 strain. F844 does not contain the influenza A(H1N1)pdm09 M gene seen in recent A(H3N2)v isolates, however both F844 and A/Indiana/10/2011 (representative A(H3N2)v virus) originate from an identical common ancestor and exhibit 93.3% identity in HA1 peptides and 88% identity in HA antigenic regions (Table 1). Both viruses show comparable identity to A/Sydney in HA1 and antigenic regions, and 27 of the 41 mutations observed in an alignment of F844, A/Indiana/10/2011 and A/Sydney HA1, are common to both F844 and A/Indiana/10/2011.
Table 1. Amino acid sequence comparison (% pairwise identity) of influenza F844, A(H3N2)v and select human H3N2 vaccine components 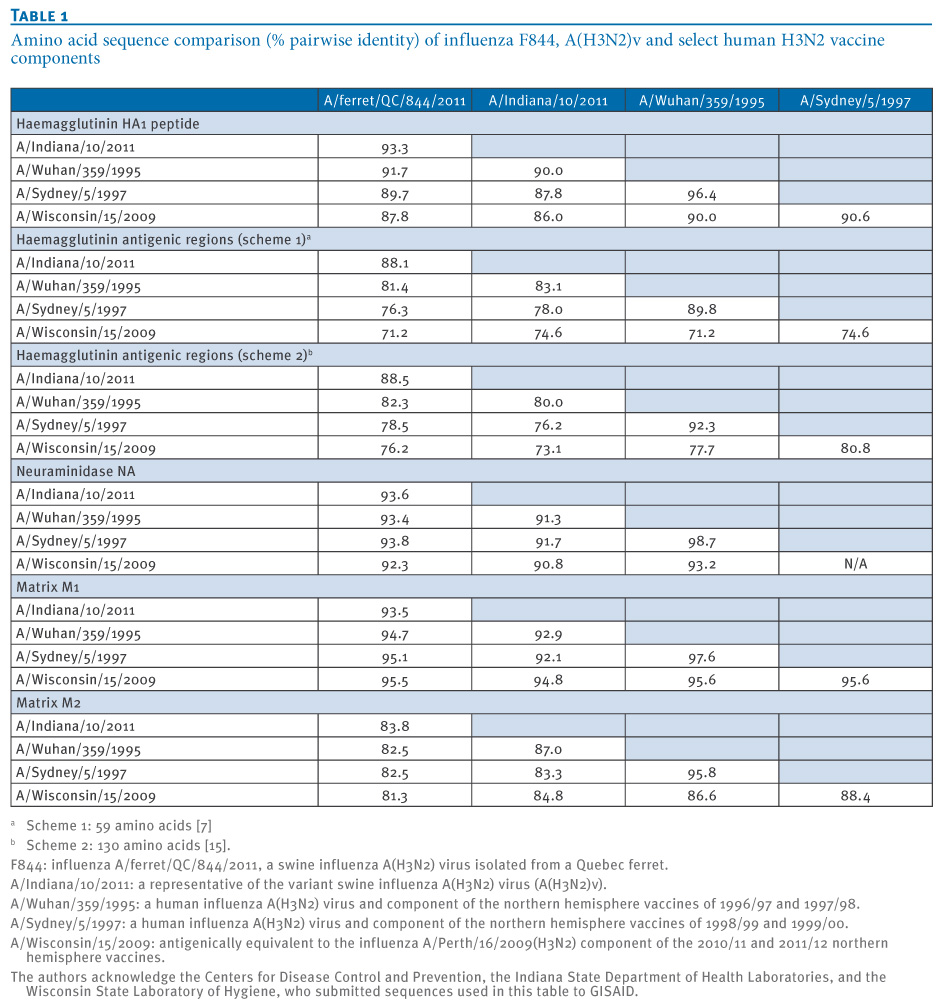
Immunogenicity and cross-reactivity
A similar proportion (ca. 35%) of children and adults showed a pre-immunisation HI titre of ≥40 to A/Wisconsin. There was substantial TIV-induced improvement in the level of antibodies to A/Wisconsin, with ca. 90% showing HI titres ≥40 post-immunisation (Table 2). For F844 and A/Sydney, only 1% and 12% of children, respectively, showed HI titres ≥40 pre-immunisation whereas approximately half of the adults showed pre-immunisation titres ≥40 to these viruses. Immunisation with the 2010/11 TIV increased antibody titres to F844 and A/Sydney only marginally in children (8% and 19% sero-converting, respectively) and adults (11% and 15% sero-converting, respectively).
Table 2. Haemagglutination inhibition antibody levels to select influenza A(H3N2) strains before and after 2010/11 trivalent inactivated influenza vaccination, Quebec, August–November 2010 (n=203)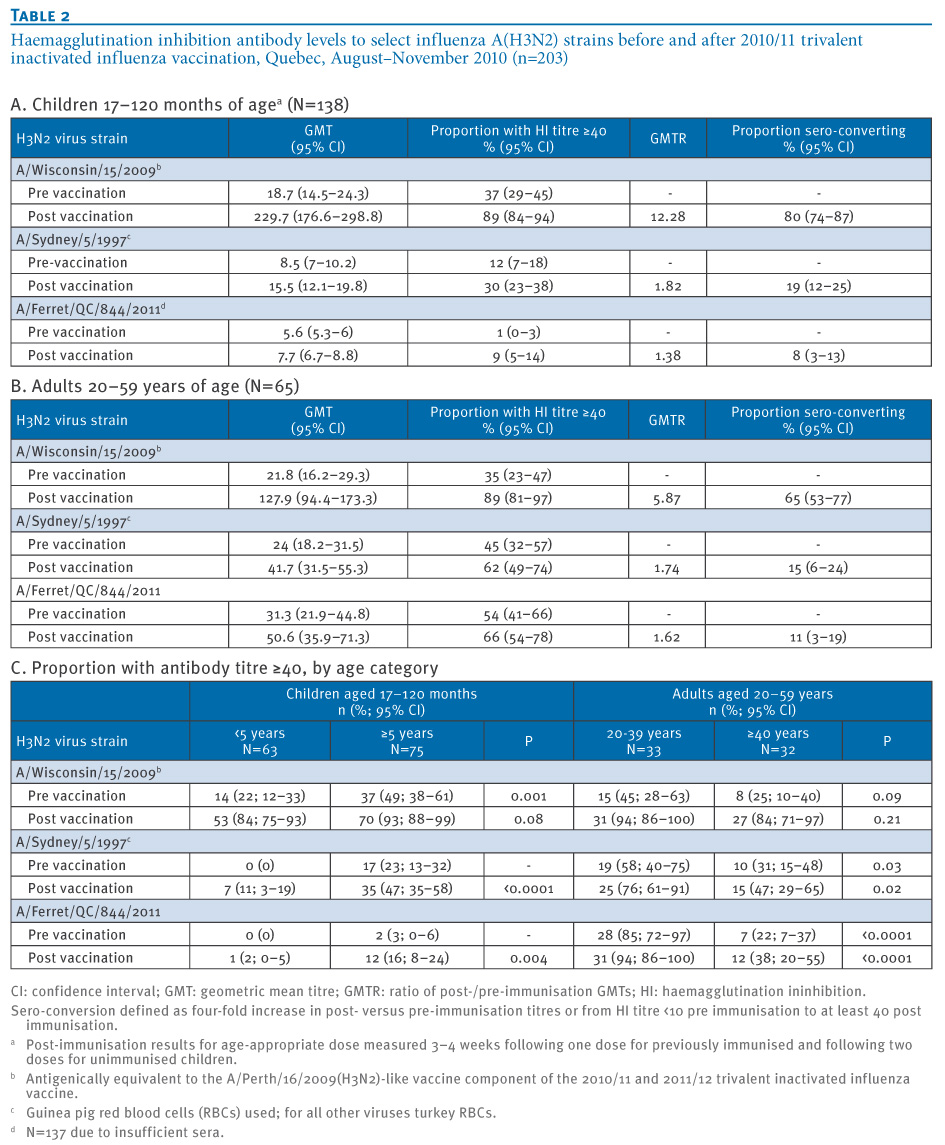
There was a strong correlation between F844 and A/Sydney titres both pre and post immunisation in children (0.74/0.76) and adults (0.68/0.76) (all p<0.001). The correlation was less strong between F844 and A/Wisconsin in both children (0.24 (p=0.005)/0.43 (p<0.001)) and adults (0.39 (p=0.001)/0.24 (p=0.05)).
On linear regression, GMTs to all three antigens increased significantly with age pre and post immunisation in the paediatric cohort; conversely in the adult cohort, GMTs decreased significantly with age for F844 and A/Sydney but not A/Wisconsin (Figure 2A/B).
Figure 2. Antibody titres pre- and post immunisation with 2010/11 trivalent inactivated influenza vaccine, by age, for influenza A/Wisconsin/15/2009, A/Sydney/5/1997 and A/ferret/QC/844/2011, Quebec, August–November 2010 (n=203)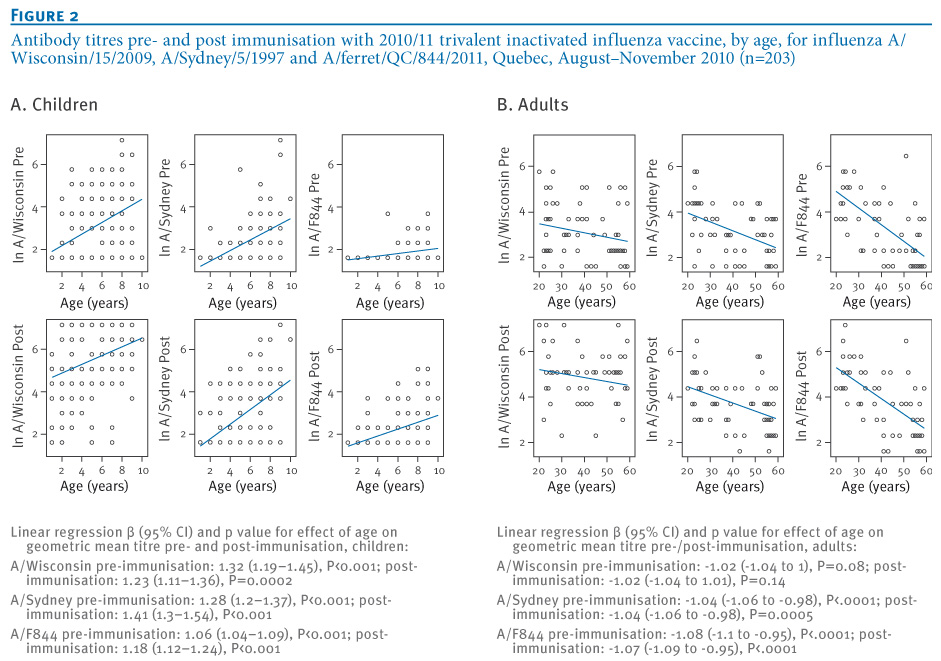
More children five years and older compared to those under five years of age had HI titres of ≥40, statistically significant for all viruses and time points except the A/Wisconsin virus at the post-immunisation collection (Table 2C). Conversely, antibody titres of ≥40 were less frequent in adults 40 years and older than in younger adults 20–39 years-old, statistically significant for F844 and A/Sydney both before and after immunisation, but not for A/Wisconsin (Table 2C).
Discussion
The intent of this investigation was to assess current vulnerability to emerging zoonotic transmissions of swine influenza H3N2 viruses. With limited early access to the specific US A(H3N2)v, we used a representative swine influenza virus locally acquired from an infected ferret to explore the likelihood of pre-existing and TIV-induced cross-reactive antibody to swine influenza A(H3N2) in children and adults. Although F844 is not a precise match to the specific A(H3N2)v, it is representative of viruses of swine-origin that have infected humans since 2005 and continue to circulate in North American pigs. F844 and A(H3N2)v showed similar pairwise identity to A/Sydney for the most relevant HA surface protein and, in combination, our findings for F844 and A/Sydney may frame major trends in cross-reactive antibody titres to swH3N2 by age. By comparing with antibodies to human viruses that are closely related but ancestral (A/Sydney) and with human strains that are antigenically distant but more recent (A/Wisconsin), we sought to better contextualise and inform age-related observations.
The limitations of our approach warrant consideration. Some laboratory variability in influenza antibody assay results is widely recognised. It should also be recognised that titres measured in HI assays, unlike microneutralisation, do not necessarily represent functional antibodies and the threshold of 40 conventionally applied as indicative of sero-protection may not predict immunity to zoonotic infections [17-21]. In general, HI is thought to overestimate cross-reactive heterologous versus homologous responses, and given that A(H3N2)v is slightly more divergent from ancestral strains in its HA1 than is F844, there may be additional reason to consider our results optimistic [18-21]. Conversely, other markers such as cell-mediated immunity may also contribute to protection but were not assessed. We have highlighted that F844 and A(H3N2)v are not precise antigenic matches and that A/Sydney is not the direct precursor of either swH3N2 virus. Differences in absolute titres or proportions by age may be expected although the major age-related trends we highlight should still apply. A specific swH3N2 virus has not yet established sustained community transmission. Further mutation, reassortment or other virus evolution is still possible and we cannot predict which swH3N2 virus may ultimately assert itself in the human population. For the purpose of ongoing risk assessment, it thus remains prudent to consider major trends rather than precise results for a particular swH3N2 virus. Finally, the small sample size, limited age categories and geographic representation of included sera must also be taken into account in interpreting our findings.
With these caveats in mind, our findings present several signals worthy of further investigation. Only few children younger than 10 years had cross-reactive antibodies to swH3N2 before or after immunisation, and when present, these antibodies were at low levels. We found more adults with cross-reactive antibody titres ≥40 against swH3N2, although GMTs were still not very high. The 2010/11 TIV only marginally increased cross-reactive antibody levels in both children and adults. Given that the same vaccine components have been used in the 2011/12 TIV, these 2010/11 findings will likely also apply to 2011/12 [10]. As noted above, however, the clinical implications of low-level cross-reactive HI antibodies are uncertain.
We observed greater likelihood of cross-reactive antibodies with increasing paediatric age but a paradoxical and unexpected pattern of decrease with increasing age in adults. We cannot address the level of cross-reactive antibodies in children 10 to 19 years of age or the elderly because they were not included in the original immunogenicity trials from which these sera were drawn. However, the pattern of decrease shown across young and middle-aged adults suggests that antibody titres may also be low in the elderly and this should prompt further evaluation.
The immuno-epidemiologic reasons for a pattern of declining cross-reactivity despite greater likelihood of cumulative exposure to influenza A(H3N2) viruses with adult age may be worth reflection. The closest ancestor of swH3N2 viruses, including A(H3N2)v, circulated in human populations approximately 15 years ago and is represented in our paper by A/Sydney/5/1997 [7]. Very young participants in this study would not have been exposed to these ancestral viruses in the mid- and late 1990s, and their lack of antibodies is therefore not unexpected; conversely, most adult participants should have been exposed. Allowing for the greatest likelihood of first influenza exposure and infection at pre-school or school age, higher titres to ancestral and related swH3N2 viruses 15 years later in young but not older adults may be consistent with the theory of robust and preferential recall of antibody to first-infecting viruses with subsequent and cumulative infections [22]. A similar phenomenon has been invoked to explain the higher pre-pandemic titres to the influenza A(H1N1)pdm09 virus found in the very old even decades after their priming exposure to a related but historic H1N1 virus in their childhood [23]. Following the same concept, lower titres to A/Sydney and related swH3N2 virus in middle-aged compared with younger adults may signal different original priming experiences. However, it was not the intent of this paper to elucidate immunological mechanisms; these concepts remain speculative and require specific hypothesis testing.
Overall, our results suggest broad susceptibility to swine-origin H3N2 infection in young children, consistent with early epidemiologic features of A(H3N2)v in the US. Susceptibility may also increase with age in adults. Given the recognised potential for children to amplify influenza spread in the community [24], and the greater vulnerability of older adults to severe outcomes of H3N2 infection generally [25], these signals warrant further investigation to guide ongoing risk assessment and response to emerging swH3N2 viruses. The H3N2 subtype of swine-origin influenza may show a different age-related pattern of risk in the human population compared to the H1N1 subtype that caused the 2009 pandemic and recommendations may need to be adjusted accordingly. We observed little TIV-induced improvement in cross-reactive antibodies suggesting that a specific candidate vaccine would be required in the event of further zoonotic transmission and epidemic spread of swH3N2 virus. Additional studies should explore age-related and vaccine-induced effects across a greater age and geographic span, applying multiple immunogenicity assays (HI, microneutralisation, cell-mediated immune markers) and swine-origin influenza A(H3N2) viruses, including A(H3N2)v.
See Also:
Latest articles in those days:
- The surveillance programme for avian influenza (AI) in Norwegian wildlife 2025 4 hours ago
- The surveillance programme for avian influenza (AI) in poultry in Norway 2025 4 hours ago
- Emergence of Novel Reassortant H3N2 Avian Influenza Viruses in Southern China: Genetic Complexity and Pathogenicity in Chickens and Mice 5 hours ago
- Pathological evidence of neurotropism and oculotropism in wild black-headed gulls naturally infected with H5N1 high pathogenicity avian influenza 5 hours ago
- Birth cohort effects in adults associated with influenza A(H1N1)pdm09 vaccine effectiveness 17 hours ago
[Go Top] [Close Window]